

开放科学(资源服务)标识码(OSID): 
0 引言
鱼类可以根据环境的变化以及自身需要采取不同的游泳模式,其整体可分为稳定游泳和不稳定的机动游泳,其中稳定的游泳为巡游,不稳定的机动游泳包括转弯、制动、摆尾滑行等[1]。过去大多数涉及鱼类运动的研究主要针对鱼类的稳定游泳,分析鱼体变形对推力的影响,缺乏对鱼类不稳定游泳的相关研究。摆尾-滑行作为鱼类常见不稳定游泳模式之一,反映了鱼类在极短时间内逃避敌害和躲避灾害环境的能力,对于鱼类的生存至关重要[2]。摆尾-滑行的特征为间歇性地摆尾,即动作由鱼体“加速”的摆尾阶段和鱼体“减速”的滑行阶段交替进行组成[3-4]。有学者研究认为鱼类采用摆尾-滑行的目的是发挥节能优势并提高机动性[3⇓⇓⇓-7],因此为解析草鱼幼鱼低能耗和高机动性的影响因素,需要进一步针对活鱼摆尾-滑行游泳动力学特征进行精细化研究。
长期以来,研究学者们定性分析鱼类摆尾-滑行的游泳特征,在运动学方向有较为充足的定量分析研究。Videler等[6]采用动力学模型预测鱼类在给定距离上使用摆尾-滑行可以节省50%以上的能量;Müller等[7]发现鱼类有2种摆尾模式即多摆尾(Multiple Tail-beat,MT)模式和单摆尾(Half Tail-beat,HT)模式,并探讨了鱼体大小对尾涡结构的影响。除了上述分析鱼类流场的方法之外,部分学者还使用了简化的试验和数值模型分析鱼类的摆尾-滑行。Chung[4]模拟发现了摆尾-滑行的节能优势,并将其归因于摆尾-滑行和巡游之间尾迹结构的不同。Wu等[8]测量了锦鲤摆尾阶段和滑行阶段的运动学特征,发现相同速度情况下锦鲤采用摆尾-滑行能比巡游节省45%的能量。Yang等[9]模拟了锦鲤摆尾-滑行与巡游的典型情况,从机械能的角度发现摆尾-滑行的效率低于巡游的效率。Li等[10]发现鱼类会控制摆尾阶段与滑行阶段时间的比率来维持所需的速度。Xia等[11]提出鱼类降低摆尾次数和摆尾阶段占整个周期的时间比有利于减少功率消耗。先前研究大多是采用简化的理论模型、无黏性的二维数值模拟分析鱼类的尾迹流场和能量优势,涉及鱼类摆尾-滑行的试验研究和定量的动力学特征分析较少,因此限制了现实场景中涉及活鱼研究的适用性。
本文使用二维PIV装置测量了静水环境下草鱼幼鱼摆尾-滑行游泳过程中的鱼体周身运动学和流体力学参数,重点分析了草鱼幼鱼头、中、尾部所受的流体推力和阻力随时间的变化特征,探究了不同流体压力下草鱼幼鱼摆尾-滑行不同阶段的游泳效率,相关研究成果能够为完善鲤科鱼类游泳行为的探索及加深鱼类摆尾-滑行运动机理的理解提供参考依据,并且可为机器鱼的优化设计提供理论基础。
1 材料与方法
1.1 试验对象
1.2 试验装置与方法
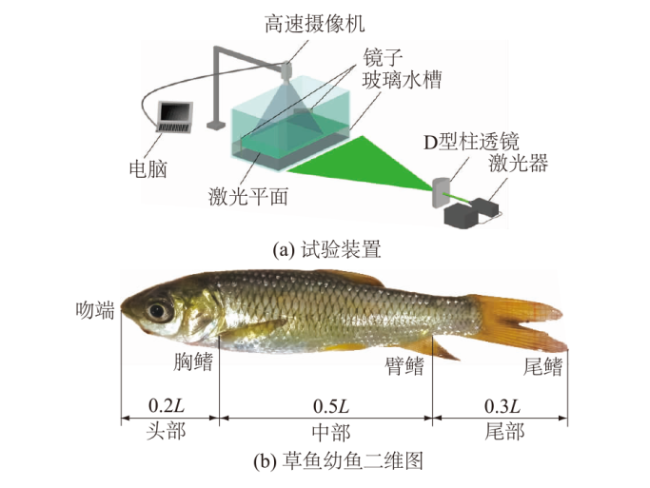
本文的分析对象为自主游动的草鱼幼鱼,为避免其他因素的干扰,本试验在黑暗、无声的环境中进行。每次试验水槽中只放入一尾鱼,每尾鱼试验15 min,每尾鱼不重复使用。水槽放水前需在水槽底部及侧壁(除了激光照射面的其他侧壁)覆盖上一层黑色防水胶带,摄像机视野内黑色背景与明亮的鱼体形成鲜明对比,以便识别鱼体-流体分界线,提取鱼体运动轮廓,减少流场测量误差。试验前将草鱼幼鱼放入水槽中,开启激光器照射试验区域,待草鱼幼鱼适应激光照射后再开始试验。为防止激光照射鱼体的另一侧出现阴影,影响后续压力点积分的计算,将2组可以调节角度的反光镜分别放置在水槽角落,调整角度以反射激光完整照亮鱼体周身的示踪粒子。
1.3 摆尾-滑行运动特征
1.4 数据提取方法
1.4.1 压力场计算
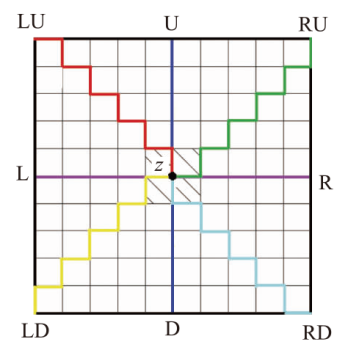
中间值周围有8条路径,每条都起源于域边界,从有限体左上角(LU)、上(U)、右上角(RU)、左(L)、右(R)、左下角(LD)、下(D)和右下角(RD)分别积分,各路径的平均值则为z点的压强,计算公式见式(1)。
式中:pU、pD、pL、pR、pLU、pLD、pRU、pRD分别表示8条积分路径上的流体压力;ΔpU、ΔpD、ΔpL、ΔpR、ΔpLU、ΔpLD、ΔpRU、ΔpRD表示相应积分路径上的压力梯度变化。
1.4.2 力学计算原理
本文结合压力场算法获取鱼体周身的流体力。当草鱼幼鱼在激光平面上进行水平运动而不采取俯仰运动,此时平面外流动小于平面流动,压力充分主导剪切力,能准确重现草鱼幼鱼不同时刻处的运动学行为和鱼体周身的流体力。此外,为了精确分析鱼体各段的压力分布,将鱼体(长度L)分成3段:吻端至胸鳍区域为头部(约0.2L),胸鳍至臀鳍的区域为中部(约0.5L),其余为尾部(约0.3L)[23]。
根据流体力学理论,鱼体-流体相互作用力F可以用式(2)计算[24]。
式中:n是法向单位矢量,方向垂直鱼体向外;p是鱼体表面流体压力;τ是黏性应力张量; 是草鱼幼鱼一个完整摆尾-滑行周期从开始到结束的时间; 是草鱼相应段的鱼体表面积。
基于流体的压力对鱼体-流体相互作用力 影响很大,当流体压力很大时,剪切力相对要小得多,忽略剪切力不会对结果产生太大的影响[25-26]。因此可以用鱼体周身的流体压力来精确估算出鱼体与流体的相互作用力。Lucas等[23]提出:若沿鱼体深度的流体压力均匀分布,则鱼体-流体相互作用力 可表示为鱼体-流体交界面处的流体压力与鱼体表面积的乘积,力的方向为鱼体-流体交界面处的法线方向。鱼体表面积的估算方法与Mchenry等[27]的相似,根据鱼体中线将鱼体等距分成200份椭圆切片,每一份的侧面积即为上述受力对应的鱼体表面积,提取每一切片的体高2a、体宽2b和切片厚度 估算出半椭圆切片的面积 。每份切片的相互作用力为
根据鱼体的前进方向,将该作用力分解成垂直于前进方向的侧向力和前进方向上的流体力(见图4),其中前进方向上的流体力Fx为
式中:θi表示每一切片上的鱼体-液体相互作用力F方向与x轴的夹角;Ai表示切片的鱼体表面积;i表示鱼体切片的数量,i=1,2,…,n。
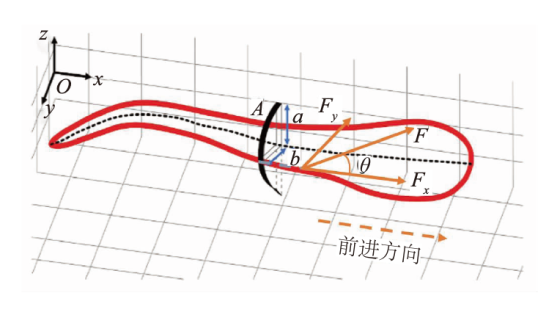
其中,与鱼体前进方向相同的力为正,定义为推力;与鱼体前进方向相反的力为负,定义为阻力,下同。
与前进方向相垂直的侧向力Fy为
鱼类瞬时时刻的游泳效率η以Borazjani等[28]提出的弗劳德效率公式表示,即:
式中: 为鱼体前进方向上的总推力; 为平均游泳速度;Plat为鱼侧向运动损耗的功率;j为侧向方向单位矢量; 为某一瞬时时刻的鱼体瞬时速度; 为草鱼每一切片的鱼体表面积。
为控制草鱼幼鱼因游泳速度的差异所导致的压力变化,鱼体两侧的流体压力被标准化为量纲的压力系数[23],即
式中 为密度。
2 结果与分析
2.1 幼鱼周身流体压力分布
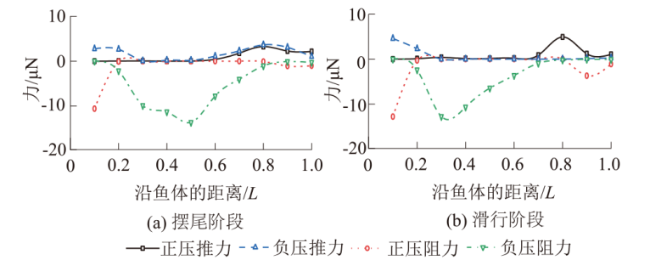
图5(a)—图5(i)显示了草鱼幼鱼摆尾-滑行阶段不同时刻的周身压力云图,其中图5(a)—图5(e)对应摆尾阶段,图5(f)—图5(i)为滑行阶段。摆尾阶段,幼鱼头部吻端始终会出现一个0.5 Pa上下波动的正压区。当鱼体向一侧摆动时,鱼体中部向相反侧弯曲,此时中部的负压值慢慢增大,而当鱼体回摆至平直状态,中部的负压区慢慢减小。摆尾阶段内,鱼尾的压力变化最大,当鱼尾向一侧弯曲,鱼体将流体向一侧推离,此时尾部弯曲内侧的正压区由0.5 Pa逐渐转变一个值为-0.45 Pa的负压区,同时弯曲的外侧会形成0.6 Pa的正压区(见图5(d))。当鱼体回摆到平直状态时,鱼体尾部的正压区、负压区均逐渐减弱,尾部正压区的最大值0.6 Pa降低至阶段末的0.3 Pa(见图5(e))。滑行阶段,鱼体保持平直状态,此时尾部的正压区却随时间慢慢增大,由阶段开始的0.3 Pa增大至结束的0.6 Pa。
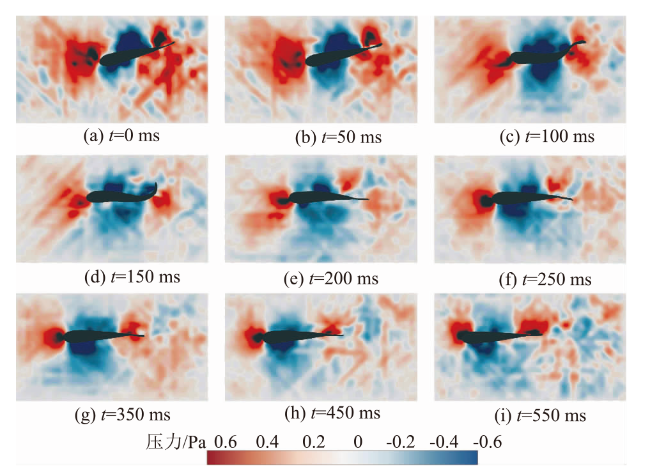
2.2 幼鱼沿鱼体方向压力系数随时间变化
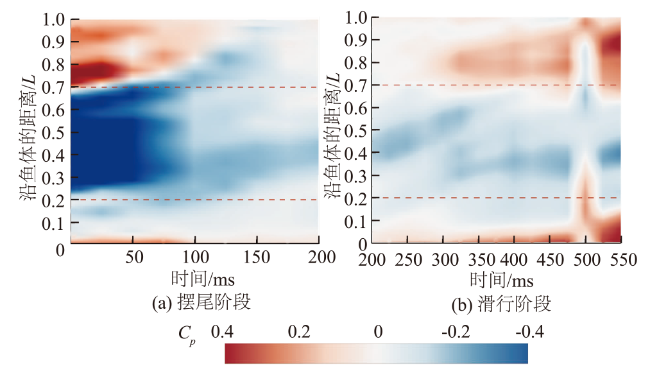
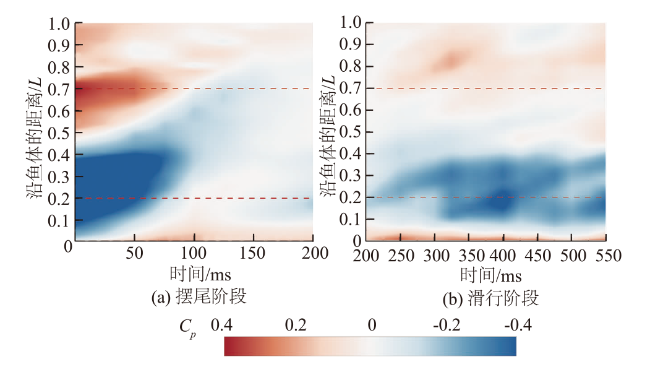
图6和图7为鱼体左、右侧轮廓沿鱼体的压力系数随时间的变化。沿鱼体方向看,鱼体各部位所受压力不同,鱼体头部(沿鱼体的距离(0,0.2L))和尾部(沿鱼体的距离(0.7L,1L))主要分布流体正压。当t=0 ms时,尾部有压力系数的峰值0.39,鱼体中部(0.2L,0.7L)两侧集中了大量的流体负压,压力系数有最小值-1。此外压力系数从负值转为正值的位置主要发生在鱼体中部靠后的地方(沿鱼体的距离(0.5L,0.7L)处)。随时间变化看,摆尾阶段(图6(a)和图7(a)),头部的压力系数由开始的0.06减小到摆尾末期的-0.1、中部由-1增大到-0.16,尾部则由0.39降低到0.1,这表明鱼体周身的流体正压和流体负压总体呈现随时间逐渐衰减的趋势;滑行阶段(图6(b)和图7(b)),鱼体中部的流体负压变化相对平稳,压力系数在-0.16上下波动,没有出现明显的增大或减小趋势,但鱼体头部和尾部的流体正压分布则随时间的推移逐渐增强,尤其尾部的压力系数,由阶段开始的0.1快速增长到结束的0.4。
2.3 推力、阻力分布特征
2.3.1 推力、阻力随时间的变化特征
整个周期内草鱼幼鱼周身推、阻力的变化曲线如图8所示。
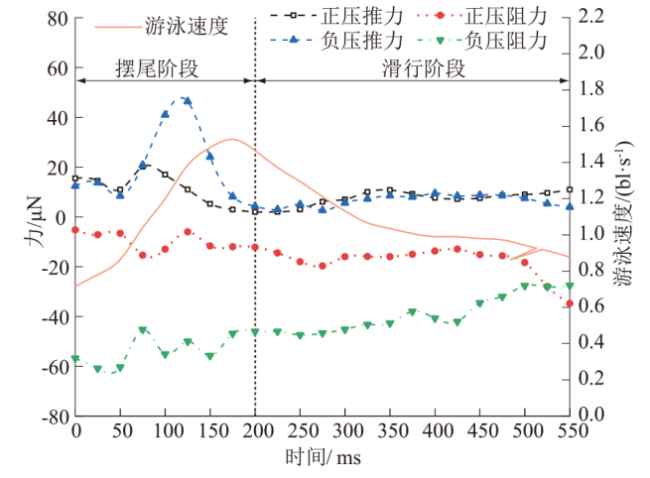
由图8可知:
(1)摆尾阶段。流体正压产生的正压推力、正压阻力均变化平稳,流体负压产生的负压推力则呈现先上升后下降的趋势,整个摆尾阶段鱼体周身流体负压产生的推力显著高于流体正压产生的推力。
(2)滑行阶段。鱼体保持笔直,鱼尾不发生摆动,压力场变化小,流体正压推力和流体负压推力分别保持在7.36 μN和6.43 μN上下。
2.3.2 推、阻力沿鱼体方向分布特征
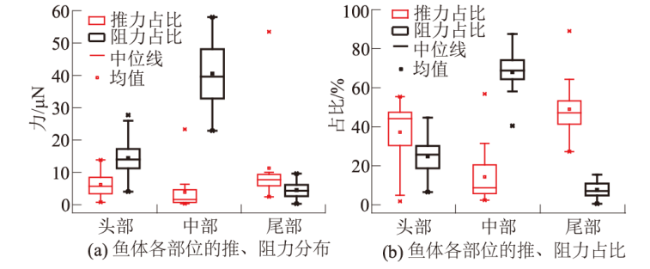
本文计算了草鱼幼鱼不同时段内推、阻力沿鱼体的分布(图9)。整个周期内,鱼体中部(0.2L,0.7L)都分布了大量的流体负压形成的阻力,而鱼体头部(0.1L,0.2L)和尾部(0.7L,1.0L)有不一样的表现。摆尾阶段,流体正、负压形成的推力主要在鱼体尾部集中分布;滑行阶段,鱼体头部有大量流体正压阻力和少量负压推力聚集。
2.4 游泳效率
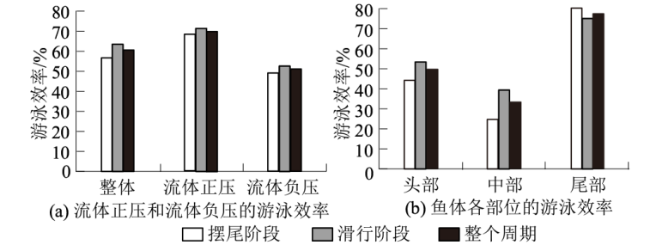
长期以来,游泳效率一直是评价鱼类高效游泳的重要指标。本文根据Borazjani等[28]提出的弗劳德效率公式计算了草鱼幼鱼基于正压、负压以及各部位处的游泳效率,其中基于正压的游泳效率表示为鱼体前进方向上基于正压推力的瞬时有效功率与基于正压的总瞬时功率的比值。整个周期内,基于流体正压的游泳效率均远高于基于流体负压的游泳效率(图11(a)),草鱼幼鱼总体平均游泳效率为60.39%±19.93%,基于流体正压的游泳效率为70.4%±21.76%,而基于流体负压的仅有51.22%±20.63%。此外,鱼体头、中、尾部的游泳效率差异显著(图11(b)),头部、中部、尾部的游泳效率分别为49.47%±24.14%、33.11%±21.5%、77.28%±16.87%,尾部的游泳效率最高,中部的游泳效率最低。
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
3 讨论
3.1 摆尾-滑行过程中鱼体的受力特征
本文通过量化草鱼幼鱼周身的流体压力分布,明确了流体正压和流体负压对鱼类摆尾-滑行的相对作用,证实了流体正压和流体负压均可以产生推力或阻力,周期内流体负压提供了占整个周期总推力55.89%的推力,但流体负压也产生了占整个周期总阻力74.44%的阻力。摆尾阶段,鱼体周身流体负压推力占总推力的38.31%,而流体正压推力占总推力的22.4%;滑行阶段,流体正压推力占总推力的21.71%,其中流体负压推力仅占17.58%。摆尾和滑行阶段流体压力分布差异反映草鱼幼鱼的不同游动形态。摆尾阶段,当鱼尾向一侧偏转时,鱼尾内侧的流体正压快速转变为流体负压,且鱼体两侧形成压力差产生了大量的推力推动鱼体前行,此时流体负压起推力的主导作用;滑行阶段,幼鱼保持笔直形态,主要依靠鱼体周身的流体正压形成推力推动自身前进。总的来说,摆尾阶段流体负压起推力主导作用,滑行阶段流体正压起推力主导作用。
另外鱼体不同部位也有不同的表现,试验结果显示:鱼体头部流体正压产生的阻力高于流体负压产生的推力,鱼体头部整体表现为阻力;鱼体中部产生高达67.82%的阻力,推力却仅有14.13%,鱼体中部总体表现为阻力,原因在于鱼体中部集中了最多的流体负压,且流体负压合力表现为阻力;鱼体尾部推力占比48.81%,大于头部和中部产生的推力,阻力却仅占7.51%,这表明尾部是推力的主要来源,结果与刘焕兴等[35]的结论一致。
3.2 鱼类不同游泳模式的动力学差异
不同流体环境下,鱼类根据自身需要采取不同的游泳策略,从而获取不同的功能优势[36]。鱼类在长途迁徙中,首要考虑因素是低能耗和高效的能量利率,而在鱼类觅食、躲避捕食者时,更多的是机动性和对其他鱼类的感知。试验发现静水环境下草鱼幼鱼会自发的采用摆尾-滑行,这可能源自于鱼类在进食、觅食或躲避捕食者时会频繁调整方向而进行不稳定的机动游泳。通过与巡游的游泳效率[29]对比发现:草鱼幼鱼自主游泳时,摆尾-滑行的平均游泳效率为60.39%±19.93%,略低于巡游的70.35%±21.84%[29],这源于速度的变化会导致效率的降低。考虑到摆尾-滑行可以发挥其方向调整和速度变化的优势,相反巡游却需要足够的时间、距离及稳定的摆尾频率,因此摆尾-滑行可以满足草鱼幼鱼面对复杂环境时兼顾高游泳效率和机动能力的综合需要,也可以为机器鱼的设计提供力学参考。
4 结论
本文利用PIV技术计算分析了静水环境下草鱼幼鱼摆尾-滑行过程中的鱼体周身流体压力分布特征,比较了草鱼幼鱼头部、中部和尾部产生的推、阻力占比和游泳效率。主要得到以下结论:
(1)草鱼幼鱼摆尾-滑行期间沿鱼体周身的流体正压、流体负压交替成为鱼体前进的主要动力:摆尾阶段,鱼体前行由流体负压提供了占总推力38.31%的主要推力;滑行阶段则是流体正压主导推动鱼体前进。
(2)针对摆尾-滑行游泳状态下,分析鱼体各部位所受推阻力时发现,48.81%的推力由尾部产生,且尾部的阻力仅占7.51%,尾部是推力的主要来源;中部产生的推力仅占14.13%,阻力却高达67.82%,鱼体中部是阻力的主要形成部位。
(3) 整个摆尾-滑行周期中草鱼幼鱼的平均游泳效率为60.39%±19.93%,其中基于流体正压的游泳效率为70.4%±21.76%,要远高于基于流体负压的游泳效率51.22%±20.63%。此外,鱼体尾部的游泳效率最高,中部的游泳效率最低。